
3.1 The History of Evolutionary Theory
Interestingly, the theory of evolution began as an attempt to explain the world under the supervision of the church. Working in the 1600s, John Ray (1627-1705) and Francis Willughby (1635-1672) are credited with some of the earliest attempts to observe and classify plant and animal life in a systematic way. As students at Trinity College at Cambridge University, they began a field later to be named natural history. Like many endeavors in the 1700s and 1800s, natural historians were employed or sponsored by the church. Charles Darwin was among them.
Darwin, Wallace, and Natural Selection
In the mid-nineteenth century, two natural historians, Charles Darwin and Alfred Russel Wallace (1823-1913), independently conceived and described the actual mechanism for evolution. Each naturalist spent time exploring the natural world on expeditions to the tropics. From 1831 to 1836, Darwin traveled around the world on H.M.S. Beagle, including stops in South America, Australia, and the southern tip of Africa. Wallace traveled to Brazil to collect insects in the Amazon rainforest from 1848 to 1852 and to the Malay Archipelago from 1854 to 1862. Darwin’s journey, like Wallace’s later journeys to the Malay Archipelago, included stops at several island chains, the last being the Galápagos Islands west of Ecuador. On these islands, Darwin observed species of organisms on different islands that were clearly similar, yet they had distinct differences. For example, the ground finches inhabiting the Galápagos Islands comprised several species with a unique beak shape (Figure 3.1).
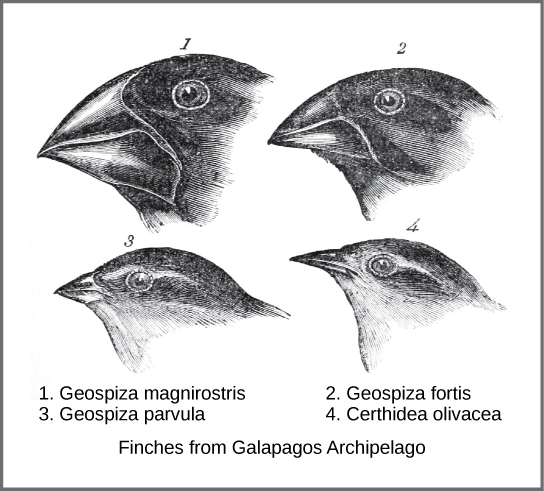
The species on the islands had a graded series of beak sizes and shapes with very small differences between the most similar. Darwin observed that these finches closely resembled another finch species on the South American mainland. He imagined that the island species might be directly related to one of the original mainland species. Upon further study, he realized that each beak shape helped the different finches acquire a specific type of food. For example, seed-eating finches had stronger, thicker beaks for breaking seeds (Fig. 3.1-1), and insect-eating finches had spear-like beaks for stabbing their prey (Fig. 3.1-4).
Wallace and Darwin both observed similar patterns in other organisms, and they independently developed the same explanation for how and why such changes could take place. Darwin called this mechanism natural selection. Natural selection is the process by which individuals with traits more suited to the current environment survive and reproduce at higher frequencies. This leads to incremental changes in a species (microevolution) that can lead to larger, species changing adaptations (macroevolution).
Natural selection, Darwin argued, was an inevitable outcome of three principles that operated in nature. He ultimately formulated the following three postulates:
- Characteristics of organisms are inherited or passed from parent to offspring.
- More offspring are produced than are able to survive, so resources for survival and reproduction are limited.
- Offspring vary among each other in regard to their characteristics and those variations are inherited.
Darwin and Wallace reasoned that offspring with inherited characteristics which allow them to best compete for limited resources will survive and have more offspring than those individuals with variations that are less able to compete. Because characteristics are inherited, these traits will be better represented in the next generation. This will lead to change in populations over generations in a process that Darwin called descent with modification.
Ultimately, natural selection leads to greater adaptation of the population to its local environment. It is the only mechanism known for adaptive evolution. Other processes such as sexual selection or domestication (seen in later chapters) are simply variations on this theme of adaptation.
In 1858, Darwin and Wallace (Figure 3.2) presented papers at the Linnean Society in London that discussed the idea of natural selection. The following year Darwin’s book, On the Origin of Species, was published. His book outlined in considerable detail his arguments for evolution by natural selection.

Gregor Mendel and His Peas
Gregor Johann Mendel (1822–1884) (Figure 3.3) was a lifelong learner, teacher, scientist, and man of faith. As a young adult, he joined the Augustinian Abbey of St. Thomas in Brno in what is now the Czech Republic. Supported by the monastery, he taught physics, botany, and natural science courses at the secondary and university levels. In 1856, he began a decade-long research pursuit involving inheritance patterns in honeybees and plants, ultimately settling on pea plants as his primary model system (a system with convenient characteristics that is used to study a specific biological phenomenon to gain understanding to be applied to other systems). In 1865, Mendel presented the results of his experiments with nearly 30,000 pea plants to the local natural history society. He demonstrated that traits are transmitted faithfully from parents to offspring in specific patterns. In 1866, he published his work, Experiments in Plant Hybridization, in the proceedings of the Natural History Society of Brünn. He was not recognized for his extraordinary scientific contributions during his lifetime; in fact, it was not until 1900 that his work was rediscovered, reproduced, and revitalized by scientists on the brink of discovering the chromosomal basis of heredity.

Mendel’s Crosses
Mendel’s seminal work was accomplished using the garden pea, Pisum sativum, to study inheritance. This species naturally self-fertilizes, meaning that pollen encounters ova within the same flower. The flower petals remain sealed tightly until pollination is completed to prevent the pollination of other plants. The result is highly inbred, or “true-breeding,” pea plants. These are plants that always produce offspring that look like the parent. By experimenting with true-breeding pea plants, Mendel avoided unpredictable traits in offspring that might occur if the plants were not true-breeding. The garden pea also grows to maturity within one season, meaning that several generations could be evaluated over a relatively short time. Mendel performed hybridizations, which involve mating two true-breeding individuals that have different traits. In the pea, which is naturally self-pollinating, this is done by manually transferring pollen from the anther of a mature pea plant of one variety to the stigma of a separate mature pea plant of the second variety.
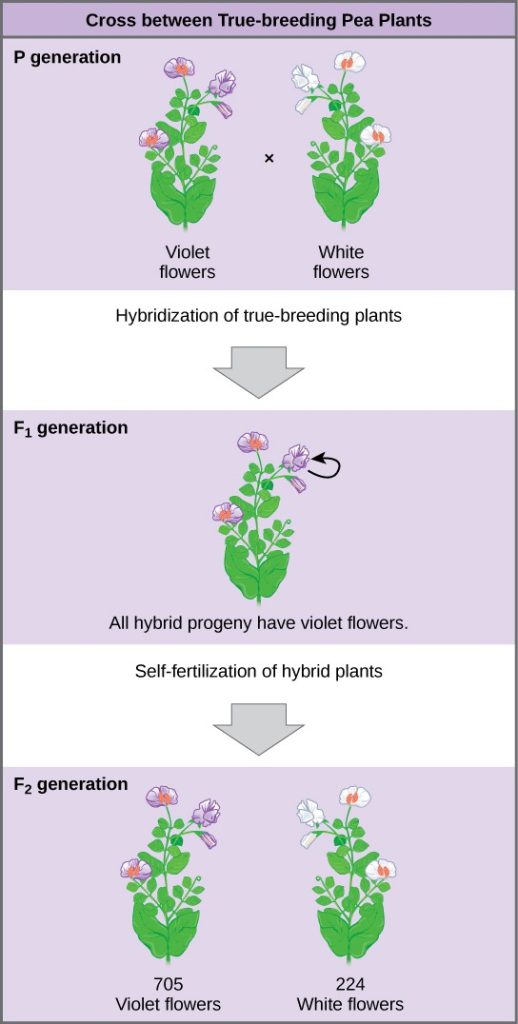
Plants used in first-generation crosses were called P, or parental generation, plants (Figure 3.4). Mendel collected the seeds produced by the P plants that resulted from each cross and grew them the following season. These offspring were called the F1, or the first filial (filial = daughter or son), generation. Once Mendel examined the characteristics in the F1 generation of plants, he allowed them to self-fertilize naturally. He then collected and grew the seeds from the F1 plants to produce the F2, or second filial, generation. Mendel’s experiments extended beyond the F2 generation to the F3 generation, F4 generation, and so on, but it was the ratio of characteristics in the P, F1, and F2 generations that were the most intriguing and became the basis of Mendel’s postulates.
Garden Pea Characteristics Revealed the Basics of Heredity
In his 1865 publication, Mendel reported the results of his crosses involving seven different characteristics, each with two contrasting traits. A trait is defined as a variation in the physical appearance of a heritable characteristic. The characteristics included plant height, seed texture, seed color, flower color, pea-pod size, pea-pod color, and flower position. For the characteristic of flower color, for example, the two contrasting traits were white versus violet. To fully examine each characteristic, Mendel generated large numbers of F1 and F2 plants and reported results from thousands of F2 plants.
What results did Mendel find in his crosses for flower color? First, Mendel confirmed that he was using plants that bred true for white or violet flower color. Irrespective of the number of generations that Mendel examined, all self-crossed offspring of parents with white flowers had white flowers, and all self-crossed offspring of parents with violet flowers had violet flowers. In addition, Mendel confirmed that, other than flower color, the pea plants were physically identical. This was an important check to make sure that the two varieties of pea plants only differed with respect to one trait, flower color.
Upon compiling his results for many thousands of plants, Mendel concluded that the characteristics could be divided into expressed and latent traits. He called these dominant and recessive traits, respectively. Dominant traits are those that are inherited unchanged in a hybridization. Recessive traits become latent in the offspring of a hybridization. The recessive trait does, however, reappear in the progeny of the hybrid offspring. An example of a dominant trait is the violet-colored flower trait. For this same characteristic (flower color), white-colored flowers are a recessive trait. The fact that the recessive trait reappeared in the F2 generation meant that the traits remained separate (and were not blended) in the plants of the F1 generation. Mendel proposed that this was because the plants possessed two copies of the trait for the flower color characteristic, and that each parent transmitted one of their two copies to their offspring, where they came together. Moreover, the physical observation of a dominant trait could mean that the genetic composition of the organism included two dominant versions of the characteristic, or that it included one dominant and one recessive version. Conversely, the observation of a recessive trait meant that the organism lacked any dominant versions of this characteristic.
Ultimately, Mendel’s work provided another crucial piece of the evolutionary theory puzzle – an explanation for Darwin’s first postulate. Yet, as time goes on, the theory would become increasingly defined.
the scientific study of animals or plants, especially as concerned with observation rather than experiment, and presented in popular rather than academic form. (Oxford Languages)
the process whereby organisms better adapted to their environment tend to survive and produce more offspring.
evolutionary change within a species or small group of organisms, especially over a short period.
major evolutionary change. The term applies mainly to the evolution of whole taxonomic groups over long periods of time.
Darwin's original term for evolution based on the idea that parents pass traits to their offspring and that small variations cause change in a species over time.
the process by which genetic information is passed on from parent to child.
an organism or other self-contained system used to evaluate a particular biologic activity or disease process.
traits that are present in the phenotype of an individual organism.
a trait that is present in the genotype and capable of emerging but has not yet become expressed.
a trait that will appear in the offspring if one of the parents contributes it.
a trait that is expressed only when genotype is homozygous and is suppressed by dominant traits in the phenotype.
