
9 Mendelian Genetics
Introduction
When Gregor Mendel started his experiments with garden peas he probably didn’t expect that the observations he made about flower color, seed shape, and plant height would lead to daytime television shows filled with “you ARE the father!” However, Mendel’s early work in the study of inheritance, the passing of characteristics between parents and offspring forms the foundations for the entire field of modern genetics.
During his experiments, Mendel quantified the probability of an individual inheriting a physical characteristic from its parents. The concept was not new. In fact, Mendel was actually recreating earlier breeding and cultivation experiments carried out by the English botanist Thomas Knight and others. What Mendel did differently was count the number of times different characteristics showed up in offspring over multiple generations.
Mendel began with pea plants that showed specific physical features. He called these characteristics traits. For example, the color of flowers, the shapes of seeds, and the heights of individual plants are all characteristics that Mendel studied. By self-fertilizing individual plants, Mendel was able to identify the exact traits present in both the male and female parents (since they were the same plant). Mendel grew several generations of plants in this manner until he found lineages that always produced offspring with the same traits. He called these lineages “true-breeding” plants. These became the parent (P) generation for his experiments.
Mendel bred true-breeding plants that showed the same traits and discovered that all offspring within those lineages always showed these same traits. However, when he crossed true-breeding plants that showed alternate traits, only one of the parental traits showed up in the next generation (F1, or first filial generation). He called the trait that showed up the dominant trait, and the one that disappeared, the recessive trait. When Mendel self-fertilized plants from the F1 generation (remember, they all looked dominant), ¾ of the next generation (F2, second filial generation) showed the dominant trait and ¼ showed the recessive trait. What Mendel decided was that the recessive trait had to be present in some of the dominant appearing plants, but that it must have been hidden.
Even more perplexing was that some of those F2 plants that showed the dominant trait produced only dominant third filial generation (F3) offspring, but others produced offspring that showed the recessive trait. Mendel observed that ⅓ of the dominant F2 plants were true-breeding, but ⅔ were not true-breeding. Further, all of the F2 plants that showed the recessive trait were true breeding for the recessive trait.
Through countless replications of these experiments, Mendel came up with five hypotheses. Paraphrasing, these are:
Key Takeaways of Mendelian Genetics
- Parents do not transfer traits (appearance) directly to their offspring. Instead, they transmit factors that contain information about these traits to their offspring.
- Every parent has two copies of the factor that governs each trait. These copies may contain the same information, or they might contain different information.
- Alternate forms of these factors (different information) lead to alternate traits (different appearance).
- The two copies of each factor do not affect each other. Neither factor changes the information carried by the other one.
- The presence of a factor within an individual does not necessarily mean that the information carried by that factor will appear in the individual.
Today, we know Mendel’s factors as genes, and the specific information that each gene carries (alternate forms of the factor), we call alleles. We also know that genes are specific portions of DNA found on chromosomes. Homologous chromosome pairs contain the same genes, but the alleles that each contains can differ. For example, the homologues of chromosome pair #1 may contain a gene for hair color, but that gene on the paternal homologue (the one you inherited from your father) may contain the allele for red hair, while the maternal homologue (that you received from mom) may contain the allele for black hair.
If the alleles that an individual inherited from their parents are the same on each chromosome of a homologous pair, that individual is homozygous for that gene. What we now call homozygous, Mendel called true-breeding. If the trait is dominant (like black hair) then the individual is homozygous dominant. If the trait is recessive, then the person is homozygous recessive. If the alleles for a given gene differ on each homologue, then the individual is heterozygous. This is what Mendel called not true-breeding. In heterozygous individuals, only the dominant trait shows in their appearance, but they carry alleles for both the dominant and recessive alleles. The collection of alleles that are present in a person’s genes is called their genotype. The collection of traits that a person displays (the physical expression of the alleles in their genotype) is that individual’s phenotype.
Writing a Genotype
To depict a person’s genotype in writing, we denote a gene using the first letter of the most common allele (also called the wild-type) within a species. In a Mendelian genetic system (1 dominant allele, 1 recessive allele), we use capital letters to depict the dominant allele and lower-case for the recessive. In our hair color example above, if red is the most prevalent hair color expressed by a species, but black hair is dominant, a homozygous dominant individual would have a genotype of RR and a phenotype of black hair. A homozygous recessive individual would have a rr genotype and a red hair phenotype. A heterozygote would have a genotype of Rr and have black hair.
If an individual has black hair, but we don’t know their genotype (remember that both homozygous and heterozygous individuals have a black hair phenotype), we denote the first allele as dominant (we know there’s at least one of these) and the second allele as a blank line or underscore. So this individual’s genotype would be written as “R__.”
Gamete Genotypes and Random Fertilization
During the formation of gametes, the homologous chromosomes of diploid germ-line cells are separated during meiosis to create haploid daughter cells. These cells are further divided and the sister chromatids are separated. In this manner, only one allele of each gene is passed to each of the haploid cells that eventually become sperm and ova. We can depict this process by writing out an individual’s genotype, then separating the alleles to form the possible gametes. A homozygous individual can produce gametes of only one allele type, but a heterozygote can produce gametes with either a dominant or a recessive allele.
| Genotype of diploid germ-line cell | AA | Aa | aa |
| Genotypes of haploid gametes produced | 100% A | 50% A, 50% a | 100% a |
We can also carry out this process for multiple genes using the mathematical shortcut called FOIL (first, outside, inside, last) to identify all possible combinations of alleles. We combine the first alleles of each gene into one gamete (first), then the first allele from gene 1 and the second from gene 2 (outside), then the second from gene 1 and the first from gene 2 (inside), and finally, the second allele from both genes (last).

During fertilization, gametes join together, and then fusion of the haploid nuclei creates a new diploid cell (zygote). We can depict this union and the possible new combinations of alleles in the zygote using a Punnett square. The gametic genotypes of each parent are listed along the top and left sides of the square. Gametes are then combined inside of each square to depict all of the possible allelic combinations in the offspring. The resulting genotype ratio depicts the probability of any child inheriting each combination of alleles. A phenotype ratio is determined by interpreting each allele combination into the physical expression shown by each offspring. Punnett squares can also be used to determine the genotype and phenotype probabilities using multiple genes. A cross involving two genes is called a dihybrid cross.
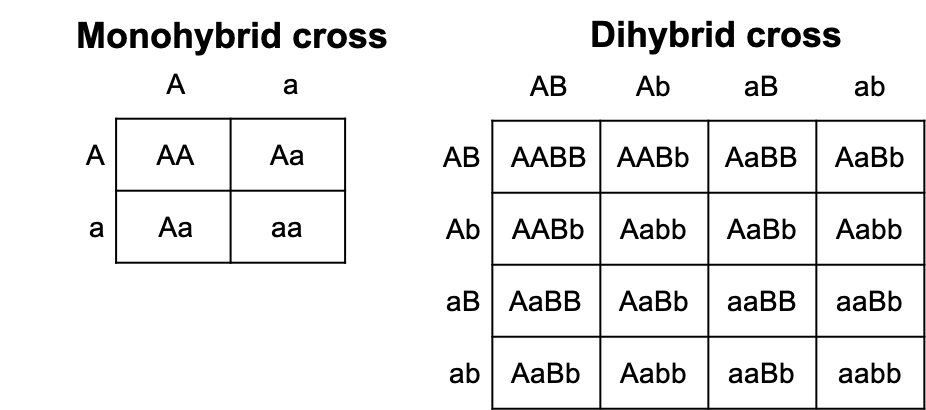
Figure 9.2 Punnett squares depict the possible genotypes of offspring by combining all of the possible genotypes from parental gametes. In a cross between parents who are heterozygous at a single gene, the offspring genotype ratio is 1 homozygous dominant: 2 heterozygous: 1 homozygous recessive. The phenotype ratio of offspring is 3:1 as the homozygous dominant and heterozygous offspring all show the dominant phenotype. In a dihybrid cross the genotype and phenotype ratios become much more complex.
Beyond Mendel: ABO Blood Type
Mendel explored traits that were characterized by single genes in a simple dominant/recessive relationship. However, most genes show more complex patterns of expression. Incomplete dominance is a pattern where the phenotype of heterozygotes is intermediate between that of homozygotes; for example, pink flowers produced by crossing red and white parents. Codominance is a pattern where two or more alleles share dominance over a recessive allele. In codominant systems, individuals who are heterozygous for the dominant alleles express both alleles in their phenotype. The human blood type is an example of a codominance system.
The genes that give us our blood types do so by encoding proteins that attach different sugars to the surface of red blood cells. These surface molecules are called antigens. An antigen is an antibody generating substance. Our bodies utilize antigens as identity markers that tell our immune system that our cells belong to us. If our immune system encounters cells that have other antigens (identity markers other than our own), we produce antibodies, proteins that bind to the antigens and mark those cells for destruction by specialized immune system cells. In blood, the reaction between antibodies in the blood plasma (liquid matrix) and foreign antigens on the surface of red blood cells causes agglutination (clotting) of the blood cells, and can cause health difficulties. Our immune system will not produce antibodies against our own antigens. However, we do produce antibodies against other antigens. In other words, if your cells produce the A antigens, your immune system will produce antibodies against the B antigens. Alternatively, if your red blood cells express the B antigens, then your immune system will produce A antibodies. But wait, it gets even more complicated than that.
The alleles that produce A and B antigens on red blood cells are codominant. This means that in someone who is heterozygous for both the A and B alleles, their red blood cells will express both A and B antigens, and they will have the AB blood type. For the A and B alleles to be codominant, they have to be dominant over something else. The something else is a recessive third allele, called the O allele. The O allele does not produce any antigen. So, an individual who is homozygous for the recessive O allele will have type O blood, and their red blood cells will not express the blood type antigens. When depicting this system on paper, we represent the codominant alleles with a capital letter I and a superscript A or B, and we represent the O allele as a lowercase i.
Following the codominance pattern, an individual can have blood type A by being homozygous for the codominant A allele (IAIA), or heterozygous for the codominant A and recessive O allele (IAi). In the same manner, they can have blood type B by being homozygous for the codominant B allele (IBIB) or heterozygous for the codominant B and recessive O allele (IBi). An individual with AB blood type will have the genotype IAIB, and someone with blood type O will have an ii genotype.
Just to complicate things further, there is another gene that affects human blood type, the so-called Rhesus factor or Rh factor. This gene was first described in the Rhesus monkey (hence the name) and is common to all primates. The positive (+) allele (dominant) produces an antigen on the surface of red blood cells, and the negative (-) allele (recessive) does not produce anything. As a result, Rh negative (Rh-) individuals produce antibodies against the Rh antigen, and Rh positive blood types (Rh+) do not produce these antibodies. The Rh gene is expressed independently of the ABO blood type gene, and therefore individuals with A, B, AB, or O blood types can be either positive or negative for the Rh factor.
You may have heard the terms “universal donor” in reference to individuals with the O negative blood type. In these individuals, the red blood cells do not have A, B or Rh antigens on them, so they will not react with the antibodies present in the blood of individuals with type A negative, B negative or AB negative. Because there are no antigens for antibodies to react with, in a blood transfusion, O negative red blood cells can be given to a person with any other blood type. In the opposite manner, individuals with type AB positive blood are considered “universal recipients.” Since these individuals do not produce antibodies against these blood antigens, they can accept red blood cells of any type in a transfusion.
Table 9.1 Blood type alleles and expression pattern in humans. Antigens are expressed on the surface of red blood cells. Antibody proteins are expressed in the blood plasma.
| Alleles | Dominance | Phenotype (Antigens expressed) |
Phenotype (Antibodies produced) |
| IA | Codominant | A antigens | B antibodies |
| IB | Codominant | B antigens | A antibodies |
| i | Recessive | none | A and B antibodies |
| R | Dominant | Rh antigen | none |
| r | Recessive | none | Rh antibodies |
Table 9.2 Genotypes and phenotypes that make up the ABO blood groups in humans. The blood type A and B alleles are codominant over the O allele. Rh factor shows a typical Mendelian dominant/recessive inheritance pattern.
| Genotype | Phenotype (Blood Type) | Genotype | Phenotype (Blood Type) |
| IAIA | A | RR | Rh+ |
| IBIB | B | Rr | Rh+ |
| IAIB | AB | rr | Rh- |
| IAi | A | ||
| IBi | B | ||
| ii | O |
Beyond Mendel: Sex Linkage
In his experiments on fruit flies (D. melanogaster), Thomas Morgan discovered a mutant allele for eye color that produced white eyes in male flies. Female flies that expressed this trait died during development. Throughout his experiments, Morgan determined that the white-eye allele was carried on the X chromosome, but not on the Y chromosome. Male fruit flies that inherited a single mutated X chromosome and a normal Y chromosome developed white eyes, but males that inherited a normal X chromosome developed normal red eyes. Females who inherited a single mutant X chromosome and a normal X chromosome developed red eyes. Thus, the white-eye allele was recessive to the red-eye allele. Morgan’s hypothesis (and subsequent confirmation) that the mutant allele was inherited strictly on the X chromosome led to an entirely new concept in the study of inheritance, that of sex-linkage.
A sex-linked trait is a characteristic that is expressed as a result of an individual’s gender because sex-linked traits are produced from genes located on the sex chromosomes. In humans (and fruit flies, as it turns out) this means the X and Y chromosomes. These traits may fall within a typical Mendelian dominant/recessive pattern of expression, or one of incomplete dominance or even codominance. For traits carried on the X chromosome, this means that males will express the inherited allele regardless of its dominance status since they only have a single X chromosome and no homologous gene on the Y chromosome. However, in females, an X-linked trait will follow the rules of expression based on its dominance pattern. If a female is heterozygous for the dominant and recessive alleles of an X-linked trait, she will express only the dominant phenotype. If the trait is incompletely dominant, she will express a phenotype intermediate between a dominant and recessive phenotype.
For traits carried on the Y chromosome (Y-linked traits), males will express any inherited allele since there is no homologous gene on the X chromosome. Females will never express these traits because they have no Y chromosome.
In this week’s lab, we will explore the genotypes and phenotypes of traits that students inherited from their parents and use a hands-on exercise to solidify the concepts of Mendelian genetics and inheritance patterns involving blood type and sex-linked traits.
Exercise 1: “Mendelian Traits”
Mendel’s work provided the foundation for our modern day understanding of how the inheritance of specific gene sequences impacts the traits of offspring. In reality, a fairly limited set of human characteristics follow classic Mendelian inheritance. Instead, many characteristics are polygenic (controlled by multiple genes) or influenced by environment and developmental processes. The table below lists easily observable traits that roughly follow Mendelian inheritance, and thus can be used to understand the inheritance of traits across generations.
- Determine your phenotype for each of these four traits (read the descriptions below the table to evaluate yourself).
- Use the allele information below to write your potential genotype(s). Remember that for dominant traits, two different genotypes can lead to the same phenotype.
- Write the potential gamete genotypes that would result from your genotype—what alleles could you pass on to offspring?
- PTC tasting: The ability to taste the chemical phenylthiocarbamide (PTC) is largely controlled by a single gene (TAS2R8) with two common alleles. Tasting the chemical is dominant to non-tasting. Use P and p to represent these alleles.
- Thiourea tasting: Like PTC, thiourea is a chemical compound that some people can taste, and others cannot. Use T and t to represent the dominant (taster) and recessive (nontaster) alleles.
- Hitchhiker’s thumb: Put your thumb out as if hitchhiking for a ride or giving a thumbs up sign. If the distal portion of your thumb points back toward your wrist, you have a hitchhiker’s thumb. Use the alleles H (hitchhiker’s thumb) and h (straight thumb).
- Interlacing fingers: fold your hands with the fingers interlaced. Which thumb is on top? Right on top is recessive to left on top. Use the alleles L (left) and l (right).
Exercise 2: Punnett squares
As a class, work through examples of Punnett squares with your instructor. For each example in your Student Workbook, start by identifying the phenotype (using the table at the end of this lab) and possible gametes of each parent. Use the parental gamete genotypes to complete the Punnett square and determine the possible offspring genotypes and phenotypes. List these and the number of each represented within the Punnett square. Finally, use the genotype and phenotype ratios to determine the probability (% chance) of producing a child with each genotype and phenotype.
Exercise 3. Mendelian virtual simulation
Purpose: The Classical Genetics Simulator (CGS) is a computer simulation of classical genetics laboratory exercises using the fruit fly, Drosophila melanogaster. The program provides you with sets of organisms with unknown patterns of inheritance and gives you the tools to design and perform experiments to discover these inheritance patterns. You will be able to mate or cross the unknown organisms and analyze your crosses in ways similar to those used by practicing geneticists and genetics counselors. CGS is not meant to replace work with real organisms. But the constraints of time, space, money and difficulty of laboratory technique make it impossible for most students to experience the excitement and challenge of such a long term real research problem. CGS can give you the chance to analyze problems that have the real surprise and real complexity that make scientific research fascinating.
What happens if you are presented with a new or previously unknown mutation in a population? How do you determine the genetic characteristics of this new mutation? The mechanism used most commonly is pedigree analysis. Pedigree analysis requires knowledge of the phenotypes of several generations of individuals and their offspring. Among humans, most genetic information comes from analysis of family trees or pedigrees. Among other organisms, for example, fruit flies, we can perform controlled crosses to determine the genetics. By analyzing the results of the matings, we can determine the genetics behind a given trait and its mode of inheritance.
